
Знаете ли вы, что основа эволюции фотосинтеза – это не разумный замысел, а любовь древних одноклеточных к халяве.
Фотосинтез – неразрешимая проблема эволюционной теории. Любой, кто изучал его, понимает: этот сложнейший биохимический путь, включающий тысячи белков, никак не поддаётся упрощению, не раскладывается на слагаемые. Каждый элемент фотосинтеза незаменим, находится в точности на своём месте и имеет неизменное строение.
Какова вероятность того, что столько генов просто возьмут и эволюционируют одновременно из ничего? Как сказал известный исследователь эволюции Стивен Блюм: «Ахаха, не смешите меня!»
Эй, фотосинтез, ну что скажешь?

Шутки шутками, но фотосинтез – один из самых “легкораскладываемых” механизмов в биологии. В нём чётко выделяются составные элементы, а также порядок, в котором они могли эволюционировать.
(1) Протонные насосы. У клетки возникают мембранные белки-насосы, которые выкачивают из неё протоны (ионы водорода).

(Фиолетовые) насосы выкачивают протоны (H+) наружу через клеточную мембрану, расходуя (красные) молекулы АТФ.
Поскольку протоны – частицы упёртые, клетке приходится тратить энергию (АТФ) на то, чтобы силой выпихивать их вон. Причём сливается энергия просто в чудовищных количествах: на работу насосов уходит до 90% всего «бюджета» горе-клетки.
Зачем всё это? Да чтоб не сдохнуть, вот зачем! Концентрация протонов в клетке (она же кислотность) – это основа основ: малейшее отклонение, и все биологические процессы встанут. Пожалуй, именно способность управлять кислотностью внутренней среды – это то, что изначально отличает живое от праха земного.
(2) Насосы на основе окисления-восстановления. Возникает новый тип насосов, который работает за счёт энергии окислительно-восстановительных реакций. Такой насос не расходует АТФ: вместо этого, он ворует высокоэнергетические электроны у молекул-доноров, растворённых в окружающей среде, и использует их энергию.

Насос отнимает электрон у (ярко-синей) молекулы-донора, пропускает через себя, и в конце отдаёт другой (бледно-фиолетовой) молекуле-акцептору.
В момент когда украденный электрон проходит через насос, он приводит его молекулярную структуру в движение своим электрическим полем, и тот буквально выталкивает протон из клетки. В конце, использованный электрон отстёгивается какой-нибудь другой молекуле (акцептору).
Поскольку АТФ больше не расходуется, с этого момента дорогостоящая прокачка протонов становится фактически бесплатной.
(3) Электрохимический градиент. Качая протоны, клетка не просто поддерживает свою кислотность – она заряжает биологический аккумулятор.

Как и полагается частицам с одинаковыми зарядами, протоны ненавидят друг друга больше всего на свете. Сила отталкивания между ними настолько огромна, что они готовы протиснуться в любую щель, лишь бы оказаться как можно дальше друг от друга.
Работа мембранных насосов создаёт разницу в концентрации заряженных частиц: протоны, выпнутые из клетки, начинают с огромной силой давить на мембрану, стремясь прорваться внутрь и выровнять концентрацию – прямо как вода, оттеснённая плотиной, всей своей массой давит на неё и рвётся на другую сторону.

Это – потенциальная энергия, которую можно использовать. К примеру, на ней работают антипортные насосы. Они пропускают протоны в клетку, и те под давлением протискиваются через каналы насосов с такой силой, что приводят молекулярный механизм насосов в действие и запускают прокачку других ионов (например калия).

Протон (H+) под давлением врывается в клетку сквозь насос и приводит в действие механизм перекачки калия (K+); АТФ при этом не расходуется.
(4) АТФ-синтаза.

На первый взгляд эта вертуха кажется сложной, но сути она состоит из двух частей: (жёлто-оранжевой) хеликазы и (синей) крутящейся фигни – скреплённых вместе белками-стабилизаторами.
Хеликаза – это моторный белок, выполняющий конкретную функцию: он садится на молекулу белка/РНК/ДНК и применяет к ней вращающий момент за счёт энергии АТФ. Другими словами, хеликаза – это приборчик для скручивания: например, с её помощью клетки развёртывают спирали ДНК.
А крутящаяся фигня – это просто колечко, воткнутое в мембрану. Вот и всё. Ничем не примечательный компонент мембранных пор и каналов.
Отвёртка, кольцо и жвачка чтобы скрепить их вместе – вот рецепт якобы самой сложной биологической машинерии в истории нашей плоской планеты.
Но как именно отвёртка и кольцо объединились в этот грациозный тандем?
Поглядим.
Сначала, ген хеликазы мутировал, что позволило ей садиться на по́ры в мембране и пропихивать через них белки за счёт энергии АТФ – что полезно для мембранного транспорта.
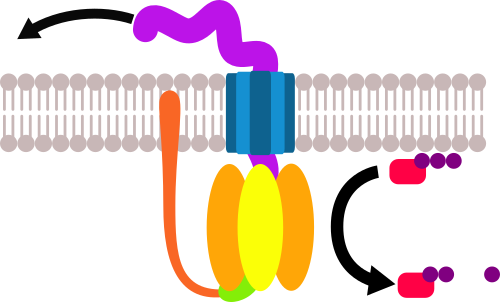
Иногда пропихиваемый белок застревал в кольце. Получалась такая вот карусель:

Это была бы самая бестолковая инновация в истории: тупо сливать АТФ на то, чтобы вертеть кольцо в мембране… если бы не одно «но»: на каждом из сегментов крутящейся фигни есть область, куда может присоединяться протон.
Поэтому, когда в стабилизирующих белках случилась мутация, давшая протонам выход к (синему) кольцу под удачным углом, получилось следующее:
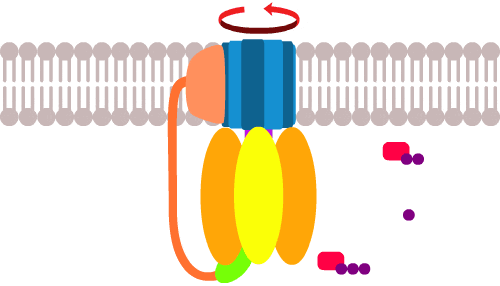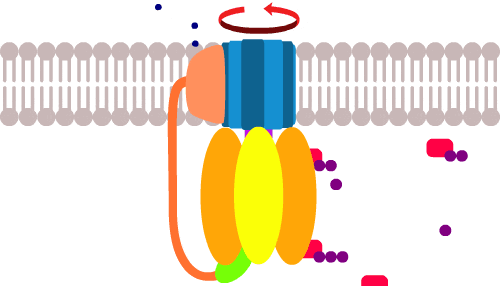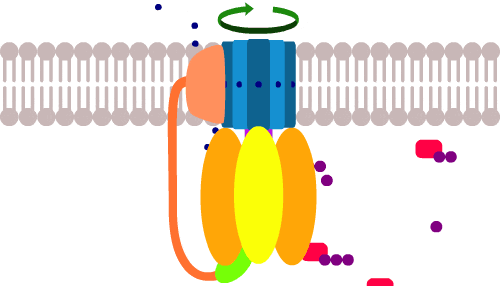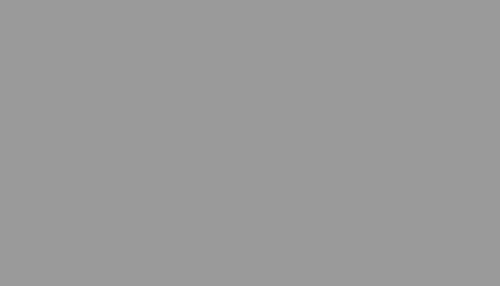
Теперь, протоны снаружи под давлением протискиваются в клетку через канал в АТФ-синтазе. По пути они цепляют и протаскивают за собой сегменты (синего) кольца и раскручивают его как мельницу в направлении, противоположном обычному.
До этого, хеликаза расщепляла АТФ чтобы вращать кольцо по часовой стрелке, но поток протонов настолько мощный, что он пересиливает энергию гидролиза АТФ, прокручивает кольцо против часовой, и по сути, запускает весь процесс в обратном порядке. Получается расщепление АТФ, только наоборот… то бишь производство АТФ.

Сказать, что эта инновация изменила жизнь клетки – значит не сказать ничего. АТФ-синтаза – это всё равно что кэшбэк 300% на любые покупки: то, что ещё позавчера нещадно сосало ресурсы из кошелька клетки стало не просто бесплатным, а наоборот преумножает ресурсы!
(здесь должна была быть реклама порядочного кэшбэка от райффайзен банка, но боюсь что ни в одном челевеческом банке таких заоблачных процентов вы даже близко не встретите)
Простор для улучшений на этом не заканчивается. При текущем раскладе электрохимический градиент нестабилен, потому что зависит от концентрации протонов вне клетки. Стоит ей нечаянно попасть в слабокислую среду, как давление протонов на мембрану тут же уйдёт в ноль, и синтез АТФ встанет.
(5) Межмембранное пространство. У клетки возникает вторая мембрана, а с ней и межмембранный тамбур, который позволяет клетке поддерживать концентрацию протонов по обе стороны от внутренней мембраны.

Отныне давление протонов на внутреннюю мембрану не зависит от кислотности окружающей среды.
(6) Насосы на основе энергии света. Переход на энергию сильных окислителей в своё время сделал прорыв, который позволил клетке списать все свои расходы на электронные доноры, тупо сидеть на мешке с неисчерпаемой халявой и ничего не делать.
Всё работало как часы.
Но проблема в том, что неисчерпаемая халява на самом деле не такая уж неисчерпаемая. Вулканы тухли, гидротермальные источники схлопывались – а вместе с тем изменился и состав океана.
Решение одно: слезать с нефтяной донорной иглы и переходить на новые источники энергии.
Так возникает новый протонный насос: вместо того, чтобы воровать чужие электроны, он поглощает фотон света – и в результате один из его собственных электронов переходит в заряженное состояние.
Проще говоря, он качает протоны за счёт энергии солнечного света:

Вот и фотосинтез подоспел. Буквально. Всего два белковых комплекса: световой насос и АТФ-синтаза (и нет, им необязательно было эволюционировать ра́зом).
Однако
Большинство фотосинтетиков на Земле используют немного другой механизм. Вернёмся к шагу №2 – насосу на основе окисления-восстановления:
Что дальше?
Всё то время пока эволюционировали АТФ синтаза и межмембранное пространство, насос тоже не стоял на месте. Под давлением отбора он постоянно оптимизировался: использовать новые доноры, качать больше протонов на каждый краденый электрон и т.п.
Наконец, насос разделяется на две части: собственно насос, и специализированный извлекатель электронов.

Отдельный (коричневый) белковый комплекс извлекает электрон у донора, после чего передаёт насосу. Перенос электрона от извлекателя к насосу осуществляет (тёмно-зелёное) вещество, растворённое в мембране.
Теперь насос только качает, а воровство электронов – это отдельная задача, выполняемая отдельным белковым комплексом. Разделение труда.
А для перегонки электронов от извлекателя к насосу клетка задействует особые вещества, растворённые в толще мембраны (например, хиноны).
Получается простейшая цепь переноса электронов.
При этом, насосу неважно, откуда берутся электроны: пока они приходят, он качает. Разделение этих функций дало клеткам возможность свободно эволюционировать новые извлекатели под разные типы доноров и оптимизировать их работу хоть до посинения, не затрагивая при этом работу насоса.
Более того, на одной мембране легко могут сосуществовать сразу несколько типов извлекателей, заточенных под разные доноры, которые шлют электроны единому насосу:

И разумеется, рано или поздно наступят тёмные времена, когда в среде больше не будет высокоэнергетических доноров в достаточном количестве.
Это – момент для выхода нового, доселе неслыханного участника электронной цепи:

(Ярко—зелёная) фотосистема поглощает свет, передаёт свой электрон насосу, а затем ворует электрон у донора, чтобы восполнить утрату.
Q: Не похоже на улучшение. Ну поглощает она какой-то там свет: электроны-то у доноров-то всё равно надо воровать?
Важно не то, надо ли воровать – а то, у кого надо воровать.
До фотосистемы работа электронной цепи зависела от сильных доноров с высокоэнергетическими электронами. Фотосистема, в отличие от обычного извлекателя, может вытаскивать электроны из любых веществ – а потом просто “подзаряжать” их светом.
Если бы высокоэнергетические электроны были золотом, то фотосистема – это философский камень. Клетка больше не зависит от доступности сильных доноров, потому что умеет превращать слабые доноры в сильные.
Растения, например, добывают электроны из воды: донора столь жалкого и ничтожного, что это даже смешно.
Кстати, электронной цепи, работающей на свету, вообще необязательно красть электроны: можно просто гонять один и тот же электрон по кругу, да то и дело его подзаряжать (так называемый циклический фотосинтез):

Всё верно: на Земле существует с десяток разных вариантов фотосинтеза – и это только то, что нам известно. Сколько их там было у вымерших видов – одному Брюсу Всемогущему известно.
Вот, например, уже ставшая классикой растительная электрон-транспортная цепь:
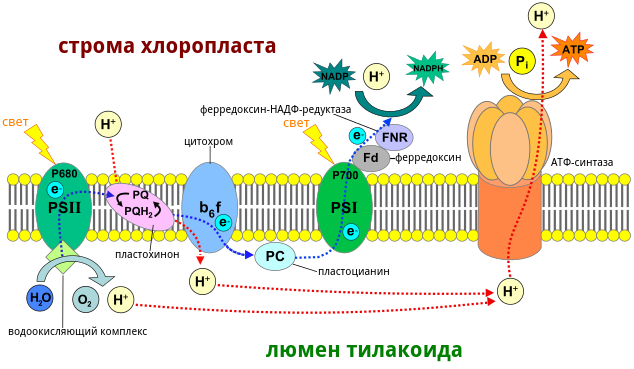
Вода как донор – такое дерьмо, что её электроны надо обдавать светом аж два раза, чтобы из них вышло что-то дельное – так что не один, а целых две фотосистемы на одну электронную цепь.
Ещё раз пройдёмся по основным этапам эволюции фотосинтеза:
-
Протонные насосы
-
Протонные насосы на основе окисления-восстановления
-
Использование энергии электрохимического градиента
-
АТФ-синтаза
-
Межмембранное пространство
-
Насосы на основе энергии света
* * *
-
Цепь транспорта электронов
-
Фотосистема
Все эти компоненты имеют независимые друг от друга функции и могли возникать по отдельности, причём практически в любом порядке (а не только в том, что приведён в статье).
Вместо заключения
Первое: нет никакого “одного фотосинтеза”. Есть десятки фотосинтетических путей разной сложности – и это лишь то, о чём нам известно.
Когда спрашивают “а разве все гены не должны эволюционировать сразу, чтобы заработал фотосинтез”, ответный вопрос: “который из?”
Второе: фотосинтез – это вообще не одна штука. Это как минимум три разных процесса, которые просто происходят одновременно:
-
Поглощение энергии света и её перевод в энергию возбуждения электронов;
-
Прокачка протонов через мембрану, нагнетение ионного градиента;
-
Синтез АТФ с использованием потенциальной энергии ионного градиента;
Все три не просто были, а остаются полезными по сей день даже по отдельности.
Q: А что с темновой фазой фотосинтеза?
Только лишний раз подтверждает главный тезис статьи: фотосинтез – это не один, а несколько процессов. По хорошему, темновые реакции (использование АТФ и НАДФН, чтобы фиксировать углерод через цикл Кальвина) вообще никак не связаны со всем описанным ранее. Это – два разных процесса, которые вполне могут существовать друг без друга.
Рекомендуемое чтиво:
Циклический фотосинтез
Про эволюцию АТФ-синтазы
Эволюция и разнообразие фотосинтетических путей
Гипотеза происхождения мембранной биоэнергетики
Ещё об эволюции фотосинтеза