
Знаете ли вы, что белый и суматранский носороги могут скрещиваться, но только через виртуального посредника?
Окажись они здесь, Аристотель и Платон сказали бы, что виды – это обособленые друг от друга категории, обладающие фиксированным набором признаков. Похожее представление закрепилось в быту: тигры – это полосатые, львы – это жёлтые с гривой. Мы смотрим на животное, видим набор признаков, и так определяем видовую принадлежность.
Вот только при всей гениальности античных философов, они не в полной мере понимали того, что мы понимаем сегодня: как на самом деле устроено биоразнообразие.
Дело в том, что биоразнообразие выглядит не вот так:

А вот так:

Не вид, и не популяция, а каждая отдельная особь обладает собственными уникальными признаками. Именно особь является базовой единицей биоразнообразия. Поэтому группу организмов – хоть стадо, хоть вид – можно описать в лучшем случае диапазонами признаков, но никак не фиксированными их значениями.
Попробуем разбить выборку на виды:

Сразу видно, что полученная систематика далека от аристотелевского идеализма:
Во-первых, некоторые виды почему-то получились “больше” (а значит, разнообразнее) других. “Расстояние” между видами тоже неоднородно: некоторые виды находятся “далеко” друг от друга (а значит, сильно отличаются), а некоторые настолько близки, что вполне сошли бы за один большой вид.
Во-вторых, границы между видами неочевидны. К примеру, можно было поделить и так:

Как доказать, что второй вариант систематики хуже или “неправильней” первого?
Но самое главное: такая систематика вообще ничем фундаментально не обоснована. Нет объективных причин считать, что граница того или иного вида должна проходить именно так, а не на сантиметр правее – и что оранизмы по разные стороны красной черты чем-то принипиально отличаются. По сути, мы нарисовали границы по принципу “получилось красиво и аккуратно”.
Что будет, если, скажем, завтра учёные встретят в лесу особи, которые не вписываются в границы? Опять всё двигать и менять?

Всё вышеизложенное – это не какие-то мысленные эксперименты, а реальные проблемы, с которыми приходится сталкиваться учёным-систематикам. К примеру, обыкновенный волк (Canis lupus) — очень разнообразный вид с огромным числом подвидов и широким размахом признаков.
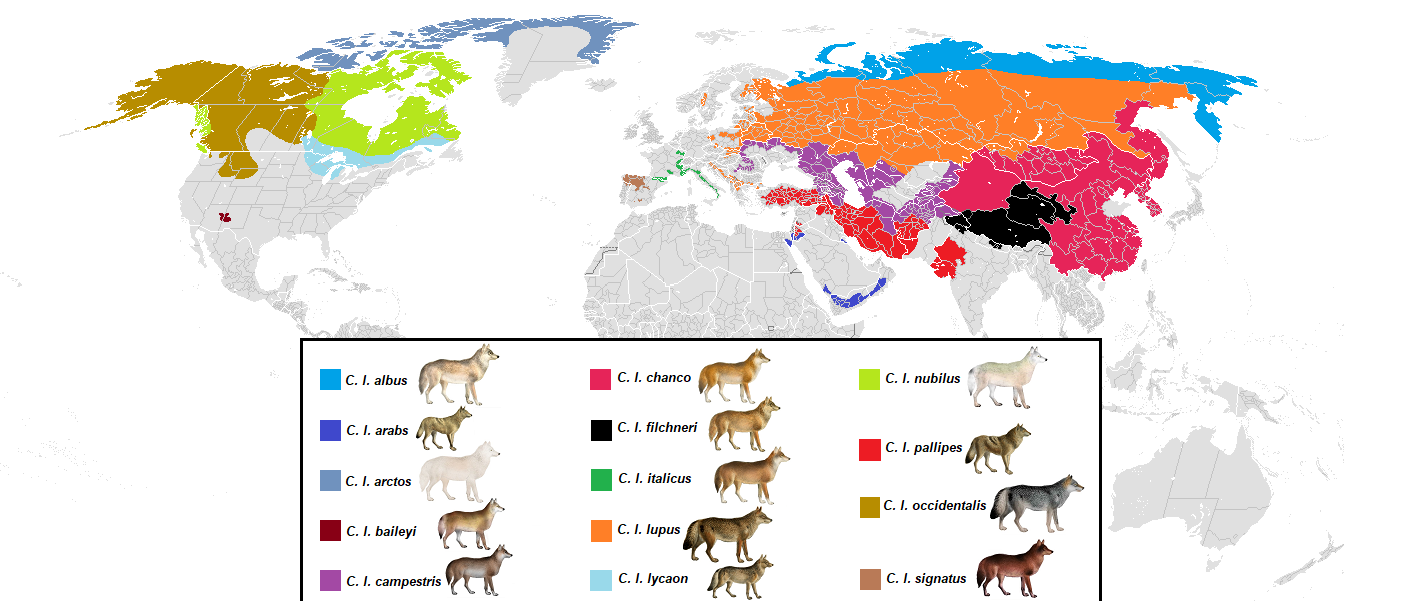
А с учётом одомашенных собак разнообразие этого вида становится просто запредельным.

А тем временем эфиопский волк (Canis simensis) — крайне однородный вид с очень узким распределением признаков.
Но при этом и обыкновенный, и эфиопский волки имеют одинаковый статус – видовой.
Есть мнение, что судить надо не по внешним признакам, а по генетическим отличиям – вот только на генетическом уровне ситуация несильно отличается. К примеру, геномы двух флоридских ланцентников (Branchiostoma floridae) могут отличаться друг от друга больше, чем геном человека отличается от генома гориллы. Выходит, что один вид ланцетников генетически более разнообразен, чем целое семейство человекообразных обезьян.

А так называемая проблема “объединителей и дробителей” – когда учёные десятилетиями не могут решить, что считать подвидом а что выносить в отдельный вид – уже давно стала обыденностью. Например, американская (Gallinula galeata) и обыкновенная (Gallinula chloropus) камышницы, которых изначально выделили в отдельные виды, потом в 1923 году объединили в один вид, и наконец в 2011 году опять поделили на два.

Gallinula chloropus слева; Gallinula galeata справа.
По большому счёту, в систематике не существует единого, универсального понятия вида: иногда это однообразная, обособленная популяция; иногда – очень разнообразная группа организмов с кучей подвидов. А иногда несколько видов и вовсе плавно перетекают друг в друга.
Отдельным пунктом обсудим страшный кошмар биологической систематики — вымершие виды. Допустим, есть выборка организмов:

Казалось бы, выборка однозначно делится на два вида. Но что будет если отобразить на графике их общих предков и все переходные формы?

Однозначно поделить такой “градиент особей” на виды невозможно в принципе. Где проходят межвидовые границы? Сколько здесь всего видов? А может всё это вообще один большой вид?

Конечно, вымерших организмов больше нет, но это не отменяет того, что они реальны, являются частью биоразнообразия, и их тоже нужно систематизировать.
Q: Разве вид — это не группа организмов, способных скрещиваться и давать плодовитое потомство?
Это — распространённое заблуждение. Такой критерий действительно есть, но он носит скорее рекомендательный характер.
Допустим, есть популяция организмов: 
Теперь выселим часть этой популяции на отдельный остров и там будем интенсивно селекционировать на протяжении десятков поколений:

В итоге получится популяция совершенно новых существ, которые не скрещиваются с исходной “зелёной” популяцией.

Выходит, мы создали новый вид. Но тогда вопрос: а к какому виду относятся особи из промежуточных популяций?

Выделить каждую из восьми популяций в отдельный вид вроде бы нельзя, потому что все они последовательно скрещиваются друг с другом. Можно попробовать сгрупировать их в два вида по четыре популяции – но тогда полученные виды будут скрещиваться друг с другом “центральными” популяциями:

Наконец, можно признать все восемь популяций одним большим видом, но тогда мы проигнорируем то, что зелёная и красная популяции сильно отличаются и не могут скрещиваться друг с другом.
Где бы мы ни провели границу между видами, особи, способные скрещиваться, в любом случае окажутся по разные стороны баррикад:
 Пожалуй, единственный способ выделить зелёную и красную популяции в два вида – это представить, что промежуточных популяций не существует. Вот только нельзя просто взять и выкинуть половину биоразнообразия из биологии.
Пожалуй, единственный способ выделить зелёную и красную популяции в два вида – это представить, что промежуточных популяций не существует. Вот только нельзя просто взять и выкинуть половину биоразнообразия из биологии.
Q: Красивая радуга из разноцветных клякс, но я не думаю что такие плавные переходы между видами существуют в реальности?
Существуют. Это называется кольцевые виды – когда две популяции не могут скрещиваться, но соединены друг с другом через цепочку из последовательно скрещивающихся популяций.
Энциклопедический пример – это цепь из популяций чаек рода Larus, обитающих на севере Евразии и Северной Америки:

(1) L. fuscus; (2) L. fuscus; (3) L. heuglini; (4) L. vegae; (5) L. vegae; (6) L. smithsonianus; (7) L. argentatus.
Популяции чаек, показанные на схеме, плавно перетекают одна в другую: к примеру, шестая (L. smithsonianus) и седьмая (L. argentatus) популяции почти не отличаются друг от друга и свободно скрещиваются, а первая и вторая популяции и вовсе относятся к одному и тому же виду (L. fuscus).
Однако в месте, где цепь смыкается в кольцо, и седьмая популяция (L. argentatus) встречается с самой первой (L. fuscus), они настолько сильно отличаются друг от друга, что не могут скрещиваться.

Larus fuscus слева, Larus argentatus справа.
Кольцевые виды – это наглядная иллюстрация видообразования: когда мы можем воочию наблюдать переходные формы между двумя видами.
В некотором смысле абсолютно все виды являются кольцевыми – просто чаще всего переходных форм нет в живых. Например, индийский (Rhinoceros sp.) и белый (Ceratotherium sp.) носороги не могут скрещиваться друг с другом, но при этом оба скорее всего смогли бы скреститься со своим общим предком.

По сути, всё живое на Земле – один большой кольцевой вид: все скрещиваются со всеми, просто некоторые делают это через “посредников”.
Если не границы, то что?
Вместо того чтобы оперировать границами, биологическая систематика основана на так называемых типовых экземплярах. Всякий раз когда учёные хотят описать или выделить новый вид, они выбирают и изымают из популяции одну конкретную особь – как правило, это наиболее репрезентативный представитель популяции с наиболее типичными и усреднёнными признаками.
Эта особь становится типовым экземпляром данного вида – образцом, на основании которого выделяется вид. Формально, видовое название привязывается не к популяции, а именно к типовому экземпляру.
Например, можно выделить четыре вида на основании следующих типовых экземпляров:
 Теперь чтобы определить, к какому виду относится любая “нетиповая” особь, не нужно пытаться вписать её в какие-либо воображаемые границы – достаточно просто сравнить её с типовыми экземплярами и определить, на который из них она похожа больше:
Теперь чтобы определить, к какому виду относится любая “нетиповая” особь, не нужно пытаться вписать её в какие-либо воображаемые границы – достаточно просто сравнить её с типовыми экземплярами и определить, на который из них она похожа больше:

Организм относится к тому виду, на типовой экземпляр которого он больше всего похож.
Точно так же систематизируется практически всё в науке и быту. Например, цвета: к жёлтому цвету относятся все оттенки, которые ближе к чистому “образцовому” жёлтому чем к чистому оранжевому или зелёному.

Плюсы такой систематики:
- Она автоматически систематизирует абсолютно все организмы, в том числе ещё не рождённые и потенциально возможные.
- Каждый вид представлен наиболее репрезентативным экземпляром, имеющим наиболее типичные для вида признаки. Это намного удобнее чем пытаться определять точные диапазоны признаков.
Вид – это множество организмов, похожих на типовой экземпляр данного вида больше, чем на типовые экземпляры других видов.
Q: Насколько сильно должны отличаться типовые экземпляры разных видов друг от друга? Можно ли, к примеру, без разбора возводить каждый встречный организм в статус типового экземпляра и выделять в отдельный вид?
Можно, но не нужно.
Сам метод систематики никак не ограничивает количество видов и степень похо́жести между ними – можно хоть братьев-близнецов выделить в два отдельных вида. Но надо понимать, что биологическая систематика – это инструмент, призванный облегчить работу и коммуникацию учёных. Поэтому существуют критерии, которым желательно следовать:
- репродуктивная обособленность,
- экологическая значимость,
- морфологические и генетические особенности,
- важность для природоохраны и оценки биоразнообразия
- и т.п.
В каждом конкретном случае решение принимает научное сообщество.
Родственные связи всё усложняют
Лучший способ удостовериться в безотказности метода – это попытаться “сломать” его: создать ситуацию, в которой он перестанет работать.
Допустим, нам надо разбить вот эту выборку на два вида:

Для этого нам надо выбрать два типовых экземпляра – самых репрезентативных представителей двух будущих видов. Логичнее всего брать откуда-то из середин двух “группировок”. Например, вот так:
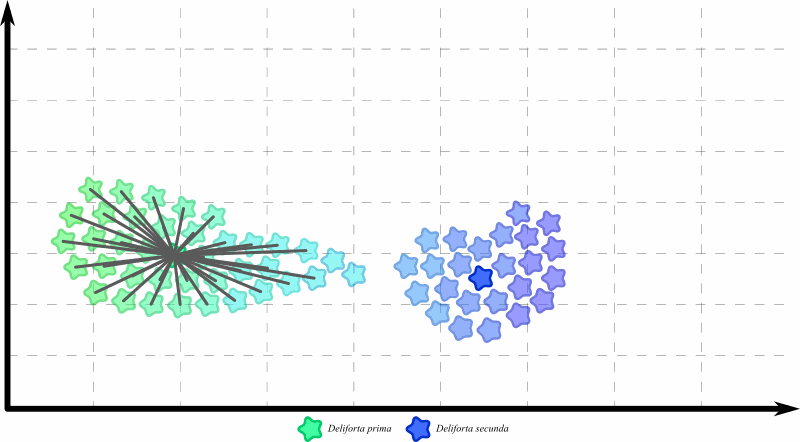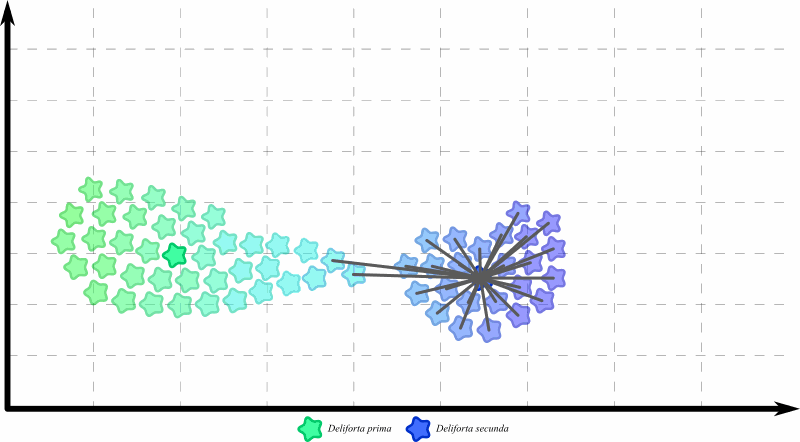
В результате, две “крайних” особи из первой кучки попали во второй вид. Выглядит не очень красиво, но справедливости ради, эти две особи действительно больше похожи на типовой экземпляр второго вида.
На этом можно было бы остановиться, если б не единственный критерий биосистематики, обязательный к соблюдению. Филогенетический, или критерий общности происхождения.
С тех пор как было установлено, что все организмы на Земле возникли в ходе эволюции и имеют общего предка, учёные сошлись на том, что эволюционное происхождение организма имеет бо́льшее значение, чем внешнее сходство: для биолога гораздо важнее знать об эволюционном происхождении органов и генов чем о сходстве их строения или функций.
Первое правило клуба: Все представители одной систематической группы должны происходить от общего предка.
Второе правило клуба: Абсолютно все представители одной систематической группы должны происходить от общего предка.
Вот почему дельфины и рыбы относятся к двум разным классам: любая систематика, которая не соблюдает филогенетический критерий, бесполезна в биологии.
Так, если сделать филогенетическую проверку (показать общих предков и восстановить общую родословную всех особей выборки), то окажется, что особи в левой и правой “кучах” образуют независимые друг от друга популяции:

Крайние особи в левой популяции не могут относиться ко второму виду, потому что они не связаны общим происхождением – и неважно, насколько они похожи. Реальное филогенетическое расстояние между ними – это не прямая линия, а вот такой заворот:

Систематика, которая учитывает эволюционное происхождение организмов, называется филогенетической. Она тоже основана на типовых экземплярах, но вместо того чтобы сравнивать признаки организмов, она сравнивает их родство. Родство определяется числом поколений, которое разделяет организмы: чем меньше поколений, тем выше родство.
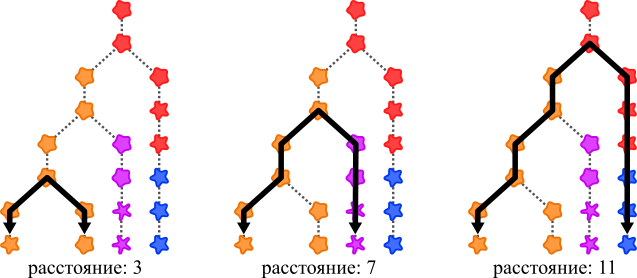
Филогенетическое расстояние между двумя организмами слева самое маленькое – следовательно, они наиболее родственны друг другу.
К слову, организмы часто бывают похожими друг на друга, имея независимое происхождение. Это называется конвергенция: когда существа из двух разных родословных приобретают схожие признаки. Как здесь:

Филогенетическое расстояние (слева) – это кратчайший путь между организмами вдоль филогенетических линий. В то время как морфологическое расстояние (справа) – это просто разница между признаками двух организмов.
Филогенетическая систематика была бы идеальным решением для биологии, если бы не одна проблема: её нереально применять на практике. Дело в том, что точную степень родства между всеми организмами невозможно установить: свечку обезьянам, жившим 4 миллиона лет назад, никто не держал (не говоря уж о тираннозаврах и трилобитах). Поэтому в большинстве случаев мы вынуждены судить о родстве по косвенным признакам: морфологическим и генетическим.
Q: А как же конвергенция?
Пример конвергенции, показанный в статье, сильно преувеличен. У реальных организмов признаков не два, а десятки тысяч. Может дельфин и сойдёт за рыбу общей формой тела, но по сотням других признаков он не имеет с ними ничего общего. Более того, не все признаки эволюционно равны: одни легко расшатываются эволюцией даже внутри одного вида, пока другие почти не меняются на протяжении сотен миллионов лет. То же самое касается и генов: некоторые последовательности ДНК могут отличаться даже внутри одной популяции, в то время как другие последовательности могут быть идентичными у целых семейств.
Правда в том, что почти всегда эволюционное родство тесно коррелирует с признаками организмов: чем два организма более родственны, тем больше они похожи – и наоборот. Во всяком случае если знать, на какие признаки смотреть. Задача систематиков и эволюционных биологов – определять “надёжные” признаки и по ним пытаться восстановить реальное филогенетическое дерево.
Филогенетическая систематика – это скорее недостижимый идеал, к которому нужно стремиться. Реальная же систематика – это лишь её приближение с помощью морфологических и генетических методов.
Итог
Идеальный вид – это все организмы, более родственные типовому экземпляру данного вида чем типовым экземплярам других видов.
Фактический вид – это все организмы, более схожие с типовым экземпляром данного вида по эволюционно значимым признакам.
F.A.Q.
Q: Что делать, если особь находится прямо между типовыми экземплярами?
Следует помнить, что систематика – это абстракция. Условность. Она существует исключительно для удобства коммуникации, и не несёт никакого вселенского смысла.
Что делать – да что угодно. Можно выделить новый, промежуточный вид, а можно приписать особь к одному из видов наугад.
Q: Если аристотелевский метод неправильный, то почему он работает? Ведь издаются же определители видов по “ключам”?
Основная причина – отсутствие переходных форм: они почти все вымерли. Из-за этого в континууме биоразнообразия есть разрывы, и это создаёт иллюзию дискретности видов.
Рекомендуемое чтиво:
Статья о принципах систематики в целом
спасибо! очень приятно и понятно читать, не могла оторваться
Спасибо за объяснение.
О трудности. Живому миру в отличие от неживого приСУЩа индиВИДуальность.